Dr Martin Juneau, M.D., FRCP
Cardiologue, directeur de l'Observatoire de la prévention de l'Institut de Cardiologie de Montréal. Professeur titulaire de clinique, Faculté de médecine de l'Université de Montréal. / Cardiologist and Director of Prevention Watch, Montreal Heart Institute. Clinical Professor, Faculty of Medicine, University of Montreal.
See all articlesOverview
- The incidence of all chronic diseases increases drastically with age and reduces healthy life expectancy.
- Several studies show, however, that the speed of aging can be greatly influenced by certain lifestyle factors, and that it may therefore be possible to delay the onset of all age-related diseases by adopting certain lifestyle habits that slow down aging.
- According to currently available data, a reduction in calorie intake, combined with regular physical activity and adequate stress management, seems to represent the optimal combination to slow down aging and thereby increase healthy life expectancy.
In Canada, as in all wealthy countries, noncommunicable diseases (cardiovascular disease, cancer, dementia, and diabetes, in particular) have become the leading causes of death and loss of healthy life years over the past century. The common feature of all these diseases is that they become much more common from middle age onwards and subsequently increase exponentially to cause a large proportion of deaths at older ages (Figure 1).
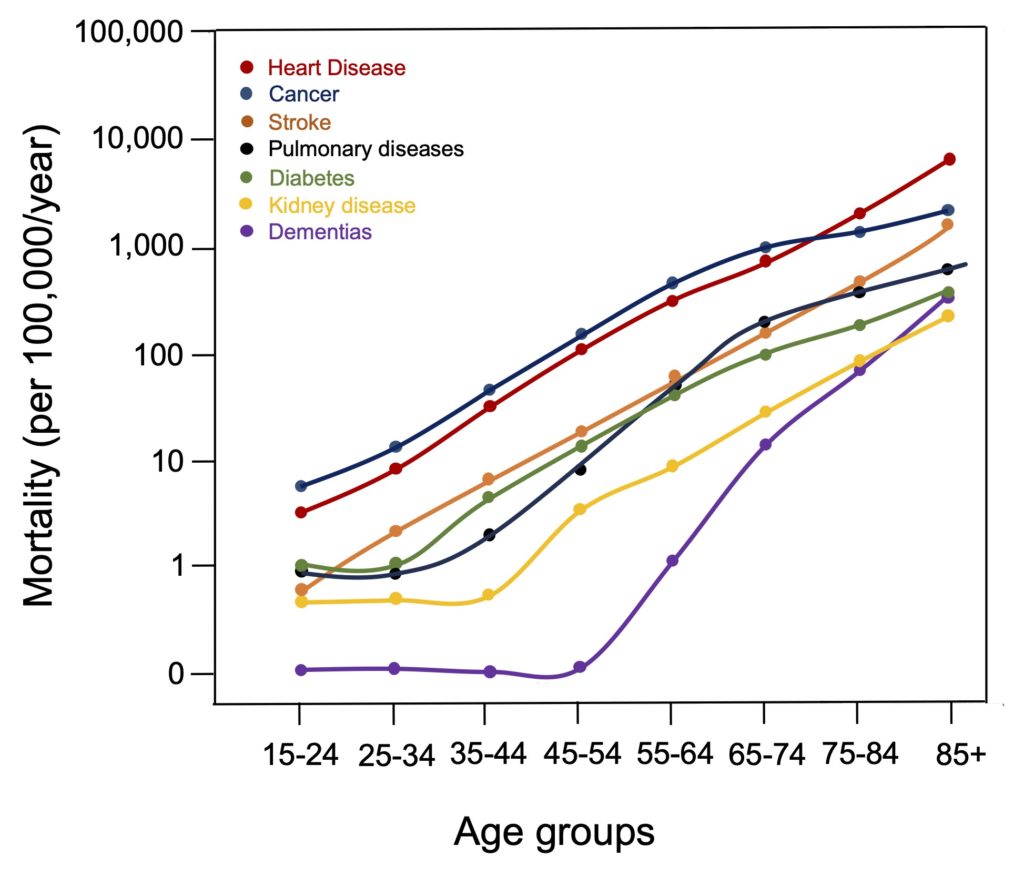
Figure 1. Increase in mortality associated with major chronic diseases during aging. Note that the number of deaths is expressed using a logarithmic scale due to the exponential increase in mortality associated with all these diseases. Data are from 1997 U.S. Vital Statistics, as presented by Miller (2002).
In addition to increasing dramatically with age, another common feature among chronic diseases is that their development is greatly influenced by a number of factors associated with lifestyle. For example, it has long been known that exposure to toxic substances (smoking, excessive alcohol consumption, pollution) and to certain metabolic (poor diet, obesity, sedentary lifestyle) or psychological (stress) imbalances increases the risk of these various diseases.
A very interesting development in research in recent years has been to show that age and lifestyle are not completely independent risk factors for chronic diseases. Several studies show that toxic aggressors and conditions such as obesity, a sedentary lifestyle, and stress accelerate cellular aging, and that this phenomenon also contributes to the increase in the incidence of chronic diseases observed during aging (Figure 2). This suggests that eliminating the factors responsible for this acceleration of cellular aging, or applying certain measures that slow down this aging process, could delay the onset of all these diseases and thus significantly increase healthy life expectancy. It is also interesting to note that in several cases of exceptional longevity, such as supercentenarians (110 years and over), the vast majority of additional years lived are in good health, with incapacitating chronic diseases only appearing late, at the end of life.
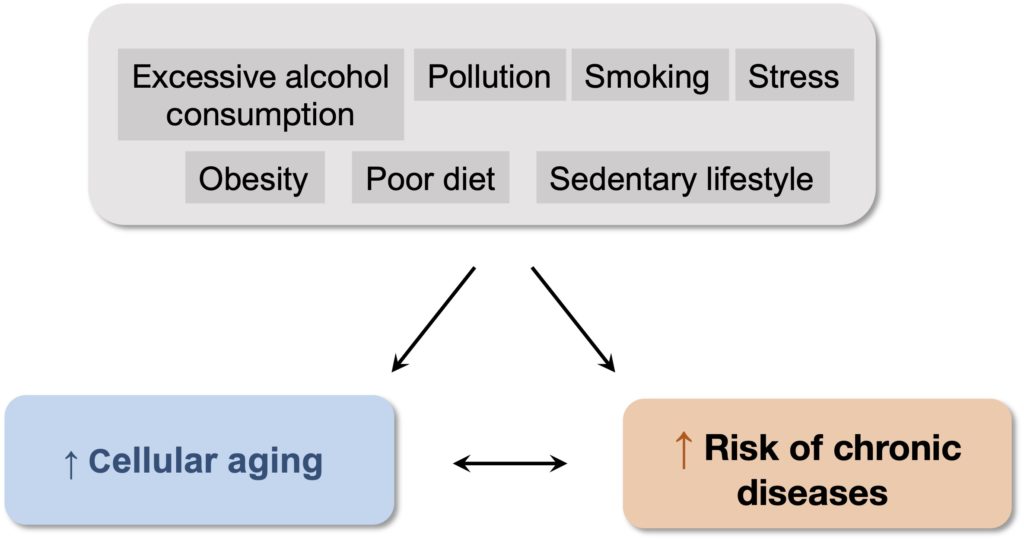
Figure 2. Contribution of cellular aging to the development of chronic diseases. All the lifestyle factors associated with an increased risk of chronic diseases cause premature aging of cells, which can amplify the impact of lifestyle on the increased incidence of these diseases that accompanies aging.
Fundamental hallmarks of aging
In the broad sense of the term, aging can be defined as a gradual decline of the various cellular systems involved in the repair and maintenance of body functions (homeostasis). In other words, aging is a process that transforms healthy young adults into less healthy older people who are at higher risk of disease and death. From a scientific point of view, research in recent years has identified 12 fundamental hallmarks that actively participate in aging, from a genetic, cellular, and organism-wide point of view (Table 1).
Table 1. The 12 fundamental hallmarks of aging. Adapted from López-Otín et al. (2022). Note that all these hallmarks are interdependent, i.e., the modulation of one process has collateral effects on others. As discussed below, the influence (positive or negative) of lifestyle on one of these hallmarks can therefore have repercussions on others and thus affect overall aging.
| Hallmarks | Description | |
|---|---|---|
| 1 | Genomic instability | The integrity and stability of the genome are constantly threatened by external stressors (chemical, physical and biological) as well as by spontaneous errors in DNA replication. Innate defence mechanisms against this damage become less effective with age, causing an accumulation of genomic damage that disrupts cellular functions. |
| 2 | Telomere shortening | Telomeres, which form “caps” at the end of chromosomes, have the function of preserving the integrity of the genome. The length of these telomeres decreases with age, leading to genomic instability and cell aging. |
| 3 | Alterations in gene expression (epigenetic modifications) | Epigenetic modifications are marks on DNA (or DNA-associated proteins, such as histones) that modulate gene expression, without altering the sequence. Aging is associated with major changes in these modifications (e.g., DNA methylation) and, consequently, in the expression of several genes essential for cell function. |
| 4 | Decline in autophagy | As the name suggests, autophagy is a process by which the cell uses its own constituents as an energy source. Autophagy is essential to eliminate dysfunctional elements that accumulate over time, both in terms of proteins and larger constituents, such as mitochondria (this is called mitophagy). The effectiveness of this cellular “renewal” activity decreases with age and contributes to the poor functioning of the cell. |
| 5 | Loss of protein balance (proteostasis) | Aging is associated with an accumulation of errors in the structure of proteins, leading to the formation of insoluble aggregates that are harmful to the cell. |
| 6 | Deregulation of nutrient measurement systems | The cell contains a system of metabolic “sensors” that specializes in detecting conditions in its environment. These sensors stimulate growth in conditions of nutrient abundance or, conversely, promote the expression of cellular defence systems when nutrients are scarcer (fasting, caloric restriction). Metabolic overheating combined with chronic exposure to high amounts of nutrients accelerates cell aging. |
| 7 | Mitochondrial dysfunction | Mitochondria are the powerhouses of the cell and are therefore essential for its functioning. With age, mitochondrial function deteriorates due to a series of changes (accumulation of mutations, deficient proteostasis) that compromise its effectiveness. |
| 8 | Cellular senescence | Senescence is a cellular response to damage that threatens its integrity. Senescent cells do not die, but rather stop dividing and subsequently acquire certain characteristics (senescence-associated secretory phenotype (SASP)) that cause local inflammation and tissue fibrosis. |
| 9 | Stem cell depletion | The role of stem cells is to renew damaged cells that can no longer perform their function. As they age, these cells are subject to the same alterations as other cells (genomic instability, loss of telomeres, etc.) and therefore gradually lose this ability to renew. |
| 10 | Altered intercellular communication | Aging is associated with deficiencies in neuronal, neuroendocrine, and hormonal signalling pathways (adrenergic, dopaminergic, insulin/IGF1, renin-angiotensin, sex hormones). |
| 11 | Chronic inflammation | Chronic inflammation is a common consequence of all the characteristics of aging (genomic instability, telomere attrition, etc.) and is manifested by an increase in the incidence of several age-related pathologies (atherosclerosis, neuroinflammation, osteoarthritis, degenerative disc disease, etc.). |
| 12 | Decline in microbiota diversity (dysbiosis) | The bacterial diversity of the microbiota is established during childhood and remains relatively stable during adulthood, but gradually declines as we age. The resulting dysbiosis contributes to the development of several pathologies such as obesity, type 2 diabetes, certain neurological disorders, cardiovascular diseases, and certain cancers. |
In addition to precisely defining the processes involved in the normal aging of the cell (and of the body as a whole), these 12 fundamental hallmarks also allow us to better understand how lifestyle can influence this aging and at the same time increase the risk of chronic diseases. Several studies show that all risk factors for chronic diseases have the common characteristic of accelerating several mechanisms involved in aging (Table 2). This negative impact is particularly striking for the two main causes of chronic diseases, obesity and smoking, but is also observed to varying degrees for all risk factors, including chemical (alcohol, pollution), physical (sedentary lifestyle), metabolic (poor diet), and psychological (stress).
Conversely, factors that have been associated in several studies with a reduction in the risk of chronic diseases, such as a reduction in calorie intake, regular exercise, and a plant-rich diet, are associated with a slowdown of several characteristics of this aging mechanism.
Table 2. Positive or negative impact of different factors associated with lifestyle on the processes involved in aging. *Alcohol consumption exceeding recommended quantities; **Western-style diet, with caloric excess and fruit and vegetable deficiency.
In other words, it is possible to modulate, positively or negatively, the speed at which aging occurs. A person’s chronological age, which reflects the linear and irreversible passage of time, is not necessarily the same as their biological age: depending on their lifestyle, they can age more or less quickly and actually have a body that is younger or older than their age. Eliminating, or at least reducing exposure to, the factors that accelerate biological aging while implementing certain interventions that slow it down, therefore represents a concrete way of preventing the development of all age-related chronic diseases and thus improving healthy life expectancy.
Eating less to live longer
One of the factors likely to slow aging that has received the most attention over the years is the reduction in food intake, whether in terms of total calories (caloric restriction) or in certain very specific elements (reduction in the intake of proteins or certain amino acids, for example). This positive effect of a restricted diet on aging may seem surprising at first glance, since it is generally believed that if something is essential to our survival (like food), a reduction in its intake should instead have negative repercussions on health. However, the exact opposite is observed: we have known for a hundred years that laboratory rodents that have access to a smaller quantity of food live longer than those that eat as much as they want. This effect of reduced calorie intake on longevity has been reproduced in several independent laboratories and in a large number of species, including yeast, nematodes (Caenorhabditis elegans), Drosophila, and non-human primates. In mammals, the increase in longevity induced by caloric restriction is correlated with a reduction in all diseases associated with aging, from cancer and cardiovascular disease to diabetes, kidney disease, and neurodegeneration.
In most cases, caloric restriction involves a reduction in all nutrients (proteins, lipids, carbohydrates) while maintaining an adequate intake of essential elements (vitamins, minerals) to avoid malnutrition. It should be noted, however, as mentioned earlier, that the positive effects of caloric restriction can be reproduced in part in certain species by specifically reducing the intake of proteins or of certain very specific amino acids such as methionine or branched-chain amino acids (BCAA: leucine, isoleucine, valine). Studies show that this protein restriction has several positive effects on metabolism, in particular glucose tolerance, insulin sensitivity, and blood levels of different classes of lipids.
The effects of long-term caloric restriction are more difficult to study in humans, but there is some evidence to suggest that it may also be associated with a reduction in the incidence of chronic diseases and premature mortality. For example, during World War I, the Danes were forced to reduce their overall food consumption for 2 years, but they did not become malnourished due to adequate intake of whole-grain cereals, vegetables and milk. This reduction in calorie intake translated into an impressive 34% decrease in mortality rates. A similar phenomenon occurred in Norway, during World War II: for a period of 4 years, the citizens of Oslo had to reduce their calorie intake by 20%, but without malnutrition (adequate intake of fresh vegetables, potatoes, fish, and whole-grain cereals). Mortality fell by 30% compared to pre-war levels, among both men and women.
Another example is the “special period” that affected Cuba during the 1990s following the fall of the Soviet Union: the economic crisis that followed caused a significant drop in the calorie intake of the population, halving the prevalence of obesity (from 14 to 7%) and causing reductions in mortality rates from diabetes, coronary heart disease, and stroke of 51%, 35%, and 20%, respectively, in the following years.
Some cultures naturally practise caloric restriction, even in the absence of tragic events that compromise the food supply. The traditional diet of the inhabitants of the Japanese island of Okinawa is undoubtedly the best example. This diet, mainly based on the consumption of plants and fish, typically contains 17% fewer calories than the Japanese average and 40% fewer calories than that of Americans. This reduced calorie intake is correlated with a premature mortality rate (60-64 years) that is half that of the Japanese in general, and a rate of centenarians 4-5 times higher than normally observed in industrialized countries. This frugal way of eating is unfortunately largely a thing of the past: the establishment of US military bases on the island after the Second World War precipitated the arrival of numerous fast-food chains and considerably modified the eating habits of the new generations. With disastrous consequences: the inhabitants of Okinawa now have the highest prevalence of obesity in Japan (38.4% among men and 25.9% among women) and one of the highest premature mortality rates in the country.
These examples clearly illustrate the positive impact associated with a reduction in calorie intake, both in terms of reduced incidence of chronic disease and healthy life expectancy. As noted in animal models, it is possible that these benefits come, at least in part, from a slowdown in aging: in the CALERIE study, for example, where participants reduced their calorie intake for 2 years by approximately 20%, there was a reduction of 2-3% in biological age, which corresponds to a 10-15% reduction in the risk of premature mortality. An improvement in metabolic health and a slowdown in aging by 2.5 years were also observed in people who had been subjected to 3 cycles of a fasting mimicking diet.
Metabolic sentinels
The mechanisms responsible for the positive effects of caloric restriction on longevity are extraordinarily complex and are still the subject of numerous studies today, both to better define the processes involved and to discover compounds that could replicate its anti-aging effects, without requiring a reduction in food intake (discussed in the next section).
It is the system involved in the metabolic response to the various nutrients in our diet (hallmark of aging #6 in Table 1) that represents the main process affected by a reduction in calorie intake. The role of this system is to allow the cell to adapt to the conditions prevailing in its environment by using 4 main metabolic sentinels that have distinct but interrelated functions (Figure 3).
- The pathway stimulated by IGF-1: IGF-1 (insulin-like growth factor) is produced by the liver in response to the secretion of growth hormone (GH) by the pituitary gland. This pathway is crucial for growth at a young age, but its overstimulation in adulthood, for example through excessive intake of calories and animal proteins, accelerates aging. It has also been observed that high levels of IGF-1 in adulthood are associated with increased risks of several chronic diseases and all-cause mortality, while conversely, people who present genetic variations that reduce the effect of growth hormone are much less affected by these diseases. Another clue to the pro-aging effect of IGF-1 comes from the differences in longevity that exist between dogs of different sizes. While very small dogs can typically live 15 years or more, larger breeds rarely reach ten years. This difference is likely due to a genetic variation present specifically in small dogs that reduces the production of IGF-1.
- AMP kinase (AMPK): The role of this sentinel is to continuously monitor the levels of ATP, the cell’s energy source. In practice, this means that energy stress (caloric restriction, for example) activates AMPK in such a way as to block processes that require ATP (e.g., growth, among others) while activating those that can generate it (drawing on sugar and fat reserves, for example). This ability to reprogram metabolism according to energy conditions makes AMPK a pivotal sentinel, capable of influencing the function of other sentinels: for example, activation of AMPK by caloric restriction results in inactivating mTOR (see below) and thus slowing down the growth of the cell.
- Sirtuins: These sentinels are enzymes (deacetylases) whose activity requires nicotinamide adenine dinucleotide (NAD) as a cofactor. Since NAD is a molecule generated by metabolism, the activity of sirtuins is intimately linked to the metabolic status of the cell. More specifically, NAD concentrations are increased under conditions of caloric restriction, and the ensuing activation of sirtuins exerts several pro-longevity effects, such as inhibition of mTOR (via activation of AMPK), improvement of mitochondrial function, reduction of inflammation, and genome stabilization. In this sense, it is interesting to note that a genetic variation that increases the function of a sirtuin seems more common in centenarians.
- mTOR (mechanistic Target Of Rapamycin): The last sentinel, and possibly the most important, is the mTOR complex. In addition to integrating information from other sentinels, mTOR also participates in the detection of energy sources, in particular the levels of proteins and certain amino acids. When these nutrients are present in high amounts, mTOR is activated and increases the synthesis of proteins, lipids, and nucleotides necessary for growth. In adulthood, growth requirements are considerably reduced and overactivation of mTOR caused by excess calories causes metabolic disturbances that can notably lead to insulin resistance. This insulin resistance contributes to the development of most chronic diseases and probably also plays a role in aging (centenarians have high sensitivity to insulin, similar to much younger individuals). One of the main effects of calorie (or protein) restriction is therefore to inactivate mTOR and avoid overheating leading to metabolic dysregulation. In other words, eating too much overactivates mTOR and accelerates aging, while reducing calorie intake inactivates mTOR and slows aging.
All this is very complicated (and again, the diagram only represents a tiny portion of the mechanisms involved!), but the point to remember is that the simultaneous and interrelated action of these sentinels allows the cell to adapt perfectly to the levels of nutrients present in the environment and to distribute resources according to its needs. During the development phase, adequate food intake activates the anabolic component (synthesis of proteins, nucleic acids, etc.) of this detection system to ensure adequate growth in children and adolescents. Once this period of growth is over, the cell’s objectives become much more focused on maintaining gains than on growth, and resources are instead directed towards maintenance processes, such as the renewal of damaged cells, the elimination of dysfunctional proteins and constituents, and DNA repair. An analogy can be made with the construction of a house: initially, the available resources are devoted mainly to the erection of the building and the purchase of the goods necessary to live there comfortably. Over time, however, the focus shifts to providing for the property’s basic needs and most resources are devoted to maintaining the existing elements to keep the house in good condition.
One of the most interesting research discoveries in recent years has been to show that it is possible to slow down the rate of aging by creating conditions that push metabolic sentinels, in particular mTOR, to promote cell maintenance rather than growth. When energy intake remains high in adulthood, for example, during the development of overweight and obesity, persistent activation of growth pathways leads to metabolic overheating that is incompatible with normal cell function and accelerates aging. Conversely, a reduced food intake, as in caloric restriction, rather allows the maintenance component to kick in, notably by reducing mTOR activity. This activates repair and recycling pathways such as autophagy, mitophagy, DNA repair, proteostasis, and stem cell regeneration (Figure 3). Since cellular aging is associated with a dysfunction of all these processes (Table 1), their activation in response to caloric restriction has the net result of slowing down this aging process.
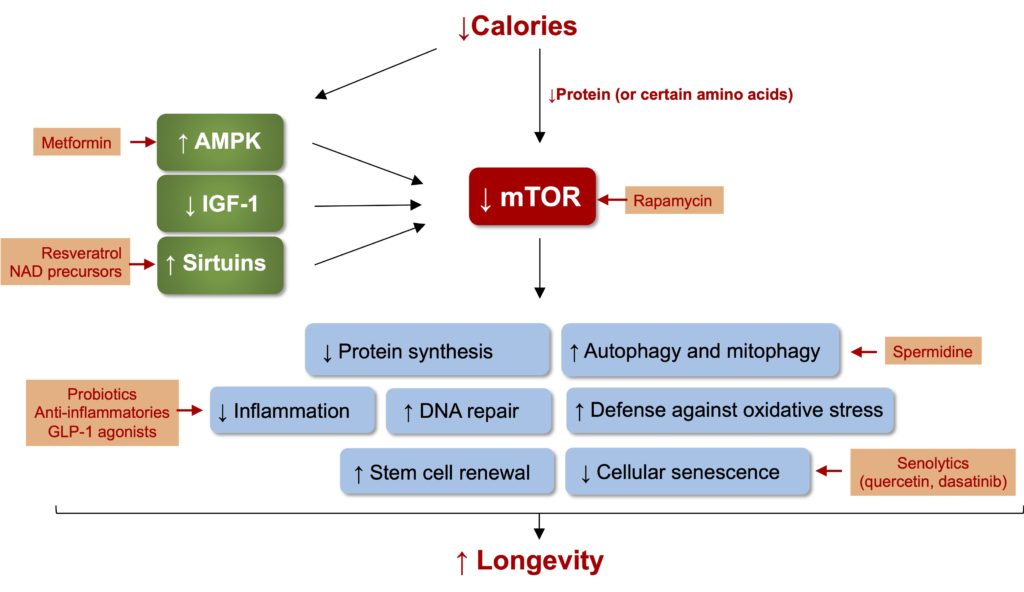
Figure 3. The central role of the mTOR complex in the increase in longevity induced by caloric restriction. The reduction in energy intake (carbohydrates, lipids, and proteins) leads to a complex response, requiring the concerted action of 4 main sentinels specialized in the detection of nutrients, namely the IGF-1 pathway, AMPK, sirtuins, and the mTOR complex. The signals coming from each of these sentinels modulate the activity of a myriad of enzymes whose action converges, for the most part, towards the inhibition of the mTOR complex. The activity of mTOR can also be directly modulated by the levels of proteins or certain amino acids such as methionine or BCAAs. Blocking the synthesis of new macromolecules leads to growth arrest and a transition to maintenance activities such as autophagy, stem cell renewal, and protein recycling (proteostasis). Since several of these processes are directly involved in cellular aging, the overall effect of caloric restriction is to slow down aging and increase longevity. The orange rectangles represent the sites of action of certain molecules currently being studied for their ability to mimic the deceleration of aging induced by caloric restriction (discussed in the following section).
Mimicking the positive effects of caloric restriction
A reduction in calorie intake remains difficult to implement, especially in the current environment where overconsumption of food is much more the norm, with no less than 1 billion people worldwide being obese. And this upward trend in excess weight shows no sign of abating, as the number of children and adolescents who suffer from obesity has quadrupled over the last 30 years, and these people are at very high risk of remaining obese in adulthood. The current dietary context, where most people have almost unlimited access to a large number of products containing significant quantities of sugar and fat that encourage overconsumption of calories, is therefore clearly not conducive to a voluntary reduction in food intake.
There are, however, alternatives: for example, instead of uniformly decreasing the amount of calories consumed each day, as in standard caloric restriction, it has been suggested that more or less prolonged periods of fasting could reproduce, at least in part, the positive effects of caloric restriction on the prevention of chronic diseases and certain characteristics of aging. Another approach, which is currently attracting a lot of interest, is to use molecules that act on certain processes influenced by caloric restriction and that contribute to increased longevity. Eight main classes of these molecules with anti-aging potential are currently being studied:
Rapamycin. In 1964, an expedition led by Montreal researchers to Easter Island (Rapa Nui) revealed, in soil samples taken from the island, the presence of a powerful antifungal produced by the bacteria Streptomyces hygroscopicus. Named rapamycin in reference to the place of its discovery, this molecule quickly distinguished itself from other antimicrobial agents due to its immunosuppressive, anticancer, and neuroprotective effects. The identification of the mTOR protein as a target of rapamycin subsequently made it possible to better understand its inhibitory effects on cell growth while highlighting the central role of this sentinel in the control of metabolism and aging (see the previous section).
In terms of aging, interest in rapamycin stems from several studies showing that this molecule significantly increases longevity in mice, yeast, the nematode C. elegans, and Drosophila. A particularly interesting effect of rapamycin is that it also appears to increase healthy lifespan, free from disease or disability, strongly suggesting that inhibition of mTOR by this molecule counters the deterioration of cellular functions that occurs during aging. In humans, on the other hand, the results obtained with rapamycin are rather mixed and remain for the moment insufficient to determine the true anti-aging potential of this molecule.
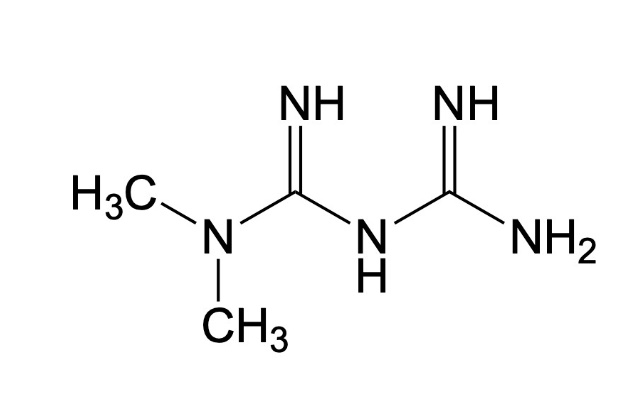
Metformin. Used for several decades for the treatment of diabetes and metabolic syndrome, metformin has more recently received attention for its prolonged effects in model systems (mice, in particular). Metformin may also improve life expectancy in humans, as diabetics treated with the molecule have been observed to live longer than non-diabetics, a surprising result given that it is well established that complications associated with diabetes are an important risk factor for premature mortality.
It seems that these positive effects on aging are mainly due to the activation of AMPK by metformin. As mentioned earlier, AMPK is a sentinel that plays a central role in the control of metabolism and influences several processes involved in aging, including mTOR activity. To further evaluate the potential of metformin to delay the development of age-related chronic diseases, a large, randomized study called TAME (Targeting Aging with MEtformin) is currently in development. The objective of the TAME study is to administer metformin (or a placebo) to people aged 65-80 years without diabetes and to compare the incidence of cardiovascular disease, cancer, and dementia between the two groups.
Resveratrol and other sirtuin activators. Certain molecules of plant origin increase the activity of sirtuins, this effect being particularly pronounced for resveratrol, a polyphenol found mainly in red wine. This activation of sirtuins is associated with a marked increase in the longevity of yeast, the nematode C. elegans, and Drosophila, and counteracts the acceleration of aging in mice subjected to a high-calorie diet.
However, the pro-longevity effects of resveratrol remain uncertain given the great variability of the results obtained by different research groups. Furthermore, a potential contribution of resveratrol to human longevity remains to be demonstrated, as no association could be observed between urinary levels of resveratrol metabolites and the risk of chronic diseases. Conflicting results have also been observed in studies using higher doses of resveratrol in supplement form, with some reporting some metabolic benefits, while others observing no significant effects. It is likely that the low bioavailability of resveratrol does not allow the molecule to reach plasma concentrations sufficient to mimic caloric restriction. Another approach used to activate sirtuins is to increase the levels of NAD+, the essential cofactor for the activity of these enzymes. For example, administering an NAD+ precursor (nicotinamide riboside) to mice was found to improve mitochondrial and stem cell function and was associated with a modest (5%) extension of the lifespan. In humans, positive metabolic effects of NAD+ precursors have been observed in some clinical studies, but, again, these benefits are not systematically observed in others. Even if caloric restriction is known to increase NAD+ concentrations (and thereby activate sirtuins), it therefore seems that pharmacological manipulation of these NAD+ levels fails to fully reproduce the positive effects of reduced food intake.
GLP-1 agonists. GLP-1 is a hormone produced in the intestines that plays an essential role in glucose metabolism by stimulating insulin secretion. Peptides that mimic the action of GLP-1 have been developed in recent years, the best known being semaglutide (Ozempic). In addition to normalizing blood sugar, these GLP-1 agonists act on the brain to activate satiety, which significantly reduces food intake (a pharmacological form of caloric restriction) and leads to weight loss. The action of these drugs on the brain also seems to slow the aging of neurons and reduce systemic inflammation.
Spermidine. Spermidine is a polyamine found throughout the living world (including microorganisms) that plays several essential roles in the maintenance of cellular functions. Plasma concentrations of spermidine can vary depending on dietary habits, as this molecule is mainly found in plant-based foods, particularly cereals, legumes, vegetables and nuts (the largest dietary source of spermidine is wheat germ).
Interest in spermidine stems from observations showing that this polyamine increases the longevity of several model systems (yeast, Drosophila, worm). Studies carried out to date on these models indicate that this positive effect is due to stimulation of autophagy, the process used by cells to get rid of defective constituents (dysfunctional mitochondria and toxic protein aggregates, for example). The impact of spermidine on human aging remains to be characterized, but studies have reported that high dietary spermidine intake is associated with a reduced incidence of cardiovascular disease and improved cognitive function.
Senolytics. Senescence is a form of permanent “pausing” of a cell in response to damage that is irreparable. Normally, senescent cells are eliminated by the immune system and replaced by new cells, but the effectiveness of this process decreases with age and the subsequent accumulation of senescent cells disrupts the body’s balance.
Several studies have focused on the identification of senolytics, i.e., molecules that can selectively eliminate senescent cells. Among the main senolytics discovered to date and the subject of particular attention are fisetin, a polyphenol found mainly in strawberries, the combination of the polyphenol quercetin and dasatinib (a drug to treat chronic myelogenous leukemia), and another polyphenol, procyanidin C1 from grape seeds.
Probiotics. The intestinal microbiota, i.e., the bacterial community that colonizes the intestine, is now recognized as a central regulator of multiple physiological processes, such as digestion and absorption of nutrients, protection against pathogens, and the production of essential metabolites (vitamins, amino acid derivatives, secondary bile acids, and short-chain fatty acids). The microbiota also sends signals to the peripheral and central nervous systems as well as to other distant organs and has an important impact on the overall maintenance of host health. It has been observed that in people who live long, healthy lives (centenarians, for example), the diversity of the microbiome tends to increase with age, suggesting that the metabolites generated by these bacteria contribute to increased longevity. This is also suggested by studies in mice where older animals that receive the microbiota of younger mice live longer. It is therefore proposed that a regular intake of probiotics could potentially contribute to maintaining intestinal microbial diversity and have positive systemic effects on the body.
Anti-inflammatory agents. Chronic inflammation increases with age, as evidenced by the increased incidence of certain inflammatory pathologies (osteoarthritis and degenerative disc disease, for example) during aging. Several studies in model systems indicate that manipulating the degree of inflammation using interventions that specifically target certain proteins involved in this process can slow aging, raising the interesting possibility that anti-inflammatory therapies may exert similar positive effects in humans.
Overall, the results obtained by these anti-aging interventions are interesting but remain at a very preliminary stage, and it is still far too early to determine their real impact on human health and longevity.
A new preventive approach
Given the current state of knowledge, most experts consider that reducing food intake represents the only concrete way to slow down aging and, thereby, reduce the risk of all age-related chronic diseases. This is an important point: it is not simply a question of living longer, but above all of increasing healthy longevity.
If (and it’s a big if) the interventions that increase longevity in animal models (caloric restriction, in particular) managed to increase human longevity in a similar way, we could expect to see life expectancies of the order of 112 years, instead of around 81 years today (Figure 4). For comparison, it has been calculated that eliminating cancer or cardiovascular disease would increase longevity by just 4 years to around 85 years. This is not so surprising, because the treatment of a given disease reduces mortality, but often fails to prevent the decline in general health and the onset of another age-related disease. So even if cures are discovered for the main age-related diseases, life expectancy remains lower than that (theoretically) obtained by slowing down aging (Figure 4).
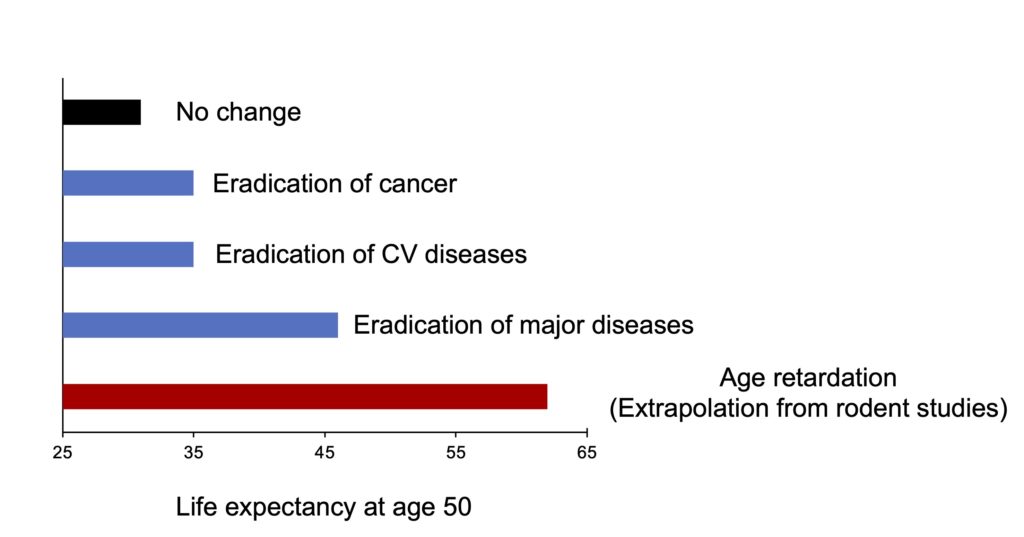
Figure 4. Variation in life expectancy at age 50 according to different hypothetical scenarios. The data represent the baseline life expectancy of a Caucasian American woman in 1985 taking into account current mortality risks (black column), or as projected under different scenarios of complete recovery from cancer or cardiovascular disease (blue columns). The red column represents the longevity that could theoretically be obtained by reproducing the slowing of aging obtained by caloric restriction in model animals (112 years). From Miller (2002).
In summary, research over the past 20 years has made remarkable progress in understanding the mechanisms involved in aging. Once considered an absolutely immutable process, it appears increasingly certain that it is possible to modulate the speed of aging and thus influence the incidence of all age-related chronic diseases. An anti-aging “pill” is certainly not for tomorrow, but we must remember that there is a broad consensus on known and proven interventions to slow aging and increase healthy life expectancy: eating fewer calories and favouring a plant-based diet, being physically active and avoiding long periods of completely sedentary time (sitting or lying down), and adequately managing stress (sleeping well, maintaining quality social connections).